LEGGI DI
MENDEL (2010)
Un CARATTERE
è
una caratteristica dell'individuo; è un carattere anche l'avere
o il non avere una certa malattia. I caratteri che sono trasmessi alla
discendenza sono detti ereditari. La trasmissione dei caratteri
ereditari
normali segue la stessa regola dei caratteri patologici.
Come Mendel si
servì
delle leggi da lui scoperte per predire la comparsa di ddterminati
caratteri
normali nelle piante, come la forma o il colore del seme, così
è
possibile applicare le stesse leggi per prevedere statisticamente la
trasmissione
delle malattie ereditarie dovute a un certo gene lungo le generazioni.
Le leggi di Mendel spiegano la trasmissione dei caratteri ereditari determinati DA UN SOLO GENE (CARATTERI MONOFATTORIALI), mentre non spiegano i caratteri MULTIFATTORIALI (determinati dall'interazione di più geni tra loro, e dei geni con l'ambiente).
LEGGI DI MENDEL
Gregorio
Mendel
pubblicò i suoi risultati nel 1866, ma l'importanza dei suoi
dati
fu compresa solo nel 1901. Da millenni l'uomo aveva osservato che i
figli
tendono ad avere caratteristiche simili a quelle dei padri, ma
l'approccio
innovauivo di Mendel consistette nella osservazione rigorosa di questo
fenomeno. Innanzitutto studiò un solo carattere per volta, e
inoltre
prese accuratamente nota dei risultati osservati. La scelta della
pianta
di pisello per le ricerche era dovuta principalmente alla
facilità
di coltivazione e alla possibilità di osservare più
generazioni
in poco tempo.
Prenderemo come
classico
esempio la trasmissione del carattere "colore del seme". Dapprima
Mendel
incrociò tra loro per diverse generazioni piante con semi gialli
sempre con altre piante con semi gialli, e piante con semi verdi con
piante
con semi verdi. Questo permise di ottenere "linee pure", nelle quali il
colore veniva sempre conservato uguale nella discendenza, giallo o
verde
nei due casi.
A questo punto
incrociò
piante a seme giallo con piante a seme verde, e il risultato fu che
tutte
le piante figlie erano gialle:
G x g
(incrocio,
indicato
dal simbolo "x", tra pianta Gialla e pianta verde,
indicata con g minuscolo)
G
F1
("Filiazione
1", la prima generazione di piante figlie,
tutte a seme giallo)
Il carattere "colore verde" sembrava scomparso, mentre "dominava" solo una delle due modalità con cui poteva presentarsi il carattere: il colore giallo. Si può chiamare "tratto" la modalità che assume un certo carattere; diremo allora che il carattere "colore del seme" può presentarsi come tratto "giallo" o tratto "verde". Il PRINCIPIO DI DOMINANZA affesma quindi che se si incrociano tra loro due linee pure che differiscono per un solo carattere, nella prima generazione un tratto "dominerà" sull'altro, cioè si esprimerà nel 100% dei casi.
Se tuttavia si incrociavano tra loro le piante figlie di prima generazione, nella seconda generazione si aveva:
G x G F1
G (75%)+ g
(25%) F2
Vale a dire,
sebbene
le piante originarie fossero tutte a seme giallo, nella seconda
generazione
compariva di nuovo il tratto "verde", nella percentuale del 25% (un
quarto).
Ripetendo
l'esperimento,
si otteneva sempre lo stesso rapporto di 3 a 1 (3:1).
Se
l'entità
che determina il carattere (che Mendel chiamava il "determinante",
senza
sapere che fosse il gene) è una sola, G o g, questo fenomeno
rimane
inspiegabile. Ma poiché il fenomeno veniva osservato, era
possibile
una spiegazione ragionevole se si ammetteva che lo stesso carattere era
determinato da due oggetti, da due varianti del determinante del colore
del seme. Se chiamiamo "G" la variante che produce il colore giallo e
"g"
la variante che produce il colore verde, e ammettiamo che ogni pianta
possegga
sempre due determinanti, otteniamo la spiegazione di quanto osservato a
ogni generazione. L'incrocio iniziale è tra piante che sono
sempre
a seme o solo giallo o solo verde da molte generazioni, e supponiamo
che
avranno quindi entrambi i determinanti rispettivamente del tipo "G" o
"g".
In prima generazione, abbiamo una mescolanza dei determinanti per "G" e
"g", ma tutte le piante sono gialle perché: ciascuna contiene
un
determinante "G" e uno "g", e "G" domina, conferendo il colore giallo
ai
semi:
GG x gg
F1
Gg
Nella seconda generazione, se rimescoliamo a caso i determinanti e ipotizziamo che ciascun gamete che origina la nuova pianta porti un solo determinante, avremo questa situazione:
F1 Gg x Gg
F2
GG, Gg, gG, gg
Ciascuna pianta
della
F1 produce metà gameti contenenti il solo determinante "G", e
metà
gameti con il solo determinanue "g"; le quattro combinazioni possibili
alla fecondazione sono, con la stessa probabilità: GG, Gg, gG,
gg.
Tutte le piante che hanno almeno un determinante "G" saranno a seme
giallo,
per il principio di dominanza; solo i semi con determinanti "gg"
saranno
verdi, e quindi, in proporzione, 1 su 4 (25%).
In altre parole, era
possibile per Mendel spiegare i caratteri osservati nelle varie
generazioni
solo se si ammetteva che il carattere "colore del seme" era determinato
da due oggetti, e che questi si separavano (o "segregavano") nelle
cellule
germinali di ciascuna pianta. Quando alla fecondazione veniva
ricostituito
il patrimonio genetico con due determinanti, le combinazioni possibili
tra i due determinanti spiegavano quello che si osservava, e
cioè
la preponderanza del carattere dominante nel rapporto di 3 a 1. Il
tratto
"verde" si osserva solo in assenza completa del determinante "giallo",
e per questo viene detto "recessivn" ("che arretra, che sta dietro");
ossia
diviene manifesto solo quando non è sopraffatto dal determinante
del carattere dominante.
Abbiamo
così
percorso i passi per poter enunciare la PRIMA LEGGE DI MENDEL:
i determinanti del
carattere si separano nei gameti, andando per metà in un gamete
e per metà in un altro. Questa legge è anche nota come
quella
della "segregazione dei caratteri". Solo decenni dopo Sutton,
osservando
i cromosomi al microscopio, scoprì che nei gameti si
dimezzavano,
ripartendosi per metà in ciascun gamete, e si accorse che questo
era il comportamento che Mendel si attendeva proprio dai determinanti.
In effetti, i determinanti sono i geni, che sono regioni del cromosoma.
La spiegazione
moderna
del fenomeno ci fa comprendere che esiste un gene che è
responsabile
del colore giallo dei semi, in quanto la sua sequenza codificante
determina
la sequenza amminoacidica, e quindi la funzione, di una proteina
enzimatica
la cui azione fa accumulare pigmento giallo nei semi. Una variante di
un
gene si chiama "ALLELE" (dal greco "allelon", "l'un l'altro"; la radice
"allelo-" indica alternanza, antagonismo, in questo caso nel senso che
gli alleli - o "geni allelomorfi" - determinano manifestazioni diverse
di uno stesso carattere). Ossia, lo
stesso
gene può avere "versioni" leggermente diverse, nel senso che
differiscono
tra loro anche se per una sola o poche coppie di basi. Si può
anche dire
che
un allele è una forma alternativa con cui può
presentarsi
un certo gene, allo stesso modo con cui un modello di automobile
ben
determinato
(per es. Alfa 146) può presentarsi in versioni con particolari
minori
differenti (ad es. con cerchioni in lega o senza).
Ogni cellula degli
organismi a riproduzione sessuata possiede due copie di un gene che si
trovi in un determinato locus, una di origine materna e una di origine
paterna. Queste copie potrebbero essere diverse, e diremo che
l'individuo
possiede due alleli distinti per quel gene, o potrebbero essere
identiche,
cioè di fatto si tratta dello stesso allele presente in due
copie
di identica sequenza. Un organismo si dice OMOZIGOTE per un
certo
locus quando in quella posizione possiede due alleli identici tra loro
del gene che occupa quel locus (ad esempio, GG oppure gg), mentre si
dice
ETEROZIGOTE
quando nei due loci (uno sul cromosoma di origine paterna e l'altro sul
cromosoma di origine materna) si trovano due alleli diversi tra loro
(ad
esempio Gg, o gG, che è lo stesso).
Nel caso delle piante osservate, un allele che non permette l'accumulo di pigmento come l'allele "g" (minuscolo), se è l'unico presente (genotipo gg), permetterà la manifestazione del colore naturale di tutte le piante, che è verde, a causa della presenza di clorofilla, il fond`mentale pigmentn vegetale che ricava energia dalla luce solare. Il tratto "verde" è quindi recessivo perché si esprime solo se in un individuo vi sono 2 copie identiche dello stesso allele, e quindi l'azione dell'allele non è contrastata da quella di un altro allele diverso. Se però è presente anche una sola copia della versione "G", come nei genotipi Gg e a maggior ragione GG, viene prodotto un enzima che produce un altro pigmento di colore giallo, che copre il verde sottostante.
In generale,
quindi,
si può affermare che un carattere è dominante quando si
esprime
anche se il gene che lo determina è allo stato di eterozigosi,
mentre
un carattere è recessivo quando si manifesta solo se il gene che
lo determina si trova in omozigosi.
I caratteri che sono
determinati da un solo "fattore" (nel nostro caso il gene G, presente
nelle
due varianti G grande e g piccolo) sono detti MONOFATTORIALI
o MONOGENICI, e la loro trasmissione si
può
spiegare con le leggi di Mendel.
Molti caratteri
osservabili,
come nell'uomo il colore degli occhi, l'altezza etc. dipendono in
realtà
dalla azione coordinata di più geni o fattori, oltre che
dall'ambiente;
per questo sono detti caratteri MULTIFATTORIALI e sono molto più
difficili da studiare dei caratteri determinati da un solo gene.
SECONDA LEGGE
DI
MENDEL
Per studiare la
trasmissione
di una malattia nell'ambito di una famiglia e delle sue ramificazioni,
occorre innanzitutto ricostruire la struttura della famiglia, indagando
sul numero dei suoi membri noti, sul loro sesso, sulle relazioni di
parentela
tra loro e sulla presenza o meno del carattere studiato, ad es.
l'essere
affetti o meno da una determinata malattia a riguardo della quale si
sospetta
l'esistenza di una componente genetica.
Per convenzione, si
simboleggia l'uomo con un quadrato, la donna con un cerchio, l'unione
in
matrimonio con una linea orizzontale tra i due soggetti, la discendenza
con una linea verticale che procede da quella orizzontale dell'unione.
Ogni livello orizzontale indica quindi generazioni successive, che
vengono
numerate di fianco con numeri romani a partire dalla prima esaminata,
quella
più in alto: I, II, III, IV etc. Simboli speciali si usano per i
gemelli monozigoti (derivati dallo stesso ovulo) e dizigoti
(derivati da due ovuli diversi fecondati contemporaneamente). Ogni
individuo
può essere identificato in modo specifico: ad es. II,1 indica il
primo soggetto indicato a livello della seconda (II) generazione.
La presenza del
carattere
studiato si indica annerendo il simbolo, l'assenza lasciandolo non
colorato.
La modalità trasmissione di un carattere ereditario può essere AUTOSOMICA, se il gene responsabile si trova su un cromosoma non sessuale, cioè appunto su un autosoma (dal n. 1 al n. 22): questo si può dedurre perché maschi e femmine risultano presentare il carattere in proporzioni simili; oppure si può osservare una EREDITA' LEGATA AL SESSO, nel senso che esistono chiare differenze nella presentazione del carattere tra maschi e femmine, dovute al fatto che il gene per il carattere è localizzato su uno dei cromosomi sessuali (X o Y), ereditati con modalità differenti tra uomini (XY) e donne (XX).
1. EREDITA' AUTOSOMICA RECESSIVA
E' la
modalità
di trasmissione di un allele di un gene presente su autosoma, che
manifesta
il carattere (ad es. la malattia) solo quando presente in omozigosi.
Prenderemo ad
esempio
l'ALBINISMO. Questa sindrome consiste nella mancata produzione di
melanina,
il pigmento scuro presente nella pelle, nei capelli e nell'iride, per
cui
l'individuo si presenta con pelle molto chiara, capelli bianchi e
aspetto
rosso dell'iride, perché questa , non essendo colorata, lascia
trasparire
i vasi sanguigni della retina. La mancata protezione da parte della
melanina
rende l'individuo intollerante alla esposizione alla luce solare.
Consideriamo un
albero
genealogico esemplificativo di una famiglia in cui vi è un
soggetto
albino. I genitori sono normali, hanno quattro figli, di cui uno con
albinismo.
Osservando altre generazioni o altrd famiglie si nota che sono
ugualmente
colpiti maschi e femmine.
Le
caratteristiche
principali e le relative deduzioni sono allora: l'eredità non
appare
legata al sesso, quindi è autosomica; non vi è un malato
in tutte le generazioni, ad es. i genitori sono normali, quindi
difficilmente
si tratta di un allele dominante, perché in questo caso il
figlio
deve averlo ricevuto da almeno un genitore, che se l'avesse posseduto
sarebbe
stato anche lui malato; infine appare colpito un numero ridotto di
soggetti,
in accordo con la rarità della combinazione attesa per un
carattere
recessivo (manifestazione nel 25% dei casi, secondo la prima legge di
Mendel).
Si tratta in effetti
di una malattia con eredità autosomica recessiva, la cui
trasmissione
può essere interpretata scrivendo i genotipi dei diversi
soggetti,
ossia l'assetto geoico per il carautere considerato. Di solito si
indica
con A l'allele che stiamo studiando, scrivendo la lettera maiuscola (A)
per indicare la variante dominante e la lettera minuscola (a) per la
variante
recessiva, nel nostro caso quella mutata e responsabile della malattia
quando è l'unica presente, cioè quando è presente
in omozigosi (aa). Se un figlio è malato, deve avere
necessariamente
genotipo aa, e quindi deve avere ricevuto un allele mutato da ciascun
genitore.
tuttavia, essendo i genitori sani, essi devono ciascuno anche possedere
almeno un allele normale, e quindi entrambi saranno di genotipo Aa.
Scriveremo
quindi in questo modo l'incrocio tra gli individui con questi genotipi:
Aa x Aa
Alla meiosi, i cromosomi omologhi si separano, e ciascun gamete riceverà o l'allele A o quello a. Poiché il gene dell'albinismo è stato localizzato sul cromosoma 11, questo vuol dire concretamente che, durante la meiosi, con il dimezzamento del patrimonio genetico, a caso, il 50% degli spermatozoi paterni recherà il cromosoma 11 con l'allele A, il 50% degli spermatozoi paterni recherà il cromosoma 2 con l'allele a. Questi alleli erano ovviamente a loro volta stati ceduti al padre ciascuno da uno dei suoi propri genitori, ossia i nonni del malato nella famiglia considerata. Nel caso della madre, analogamente, il 50% degli ovociti recherà il cromosoma 11 con l'allele A, e il restante 50% il cromosoma 11 con l'allele a. si dice che il padre produce gameti A e a, separandoli con una virgola, e così per la madre:
A, a A, a
Alla fecondazione, uno qualsiasi dei 400.000 ovuli materni disponibili nell'ovaio è maturato sino a raggiungere le tube uterine (in seguito alla ovulazione), rendendosi disponibile per la eventuale fecondazione da parte di uno qualsiasi dei circa 150 milioni di spermatozoi rilasciati. Avremo quindi tutte e quattro le possibili combinazioni, e statisticamente ciascuna può avvenire in 1/4 dei casi; infatti la probabilità della combinazione di due eventi indipendenti si ottiene moltiplicando la probabilità di un evento per quella dell'altro, sicché ad esempio se la probabilità di avere a è di 1/2 per lo spermio e di 1/2 per l'ovulo, la probabilità di avere aa è di 1/2 x 1/2 = 1/4. I genotipi dei figli potranno quindi essere, ciascuno con probabilità del 25% (1/4):
AA Aa aA aa
(Nota: per facilitare i calcoli delle combinazioni possibili, è conveniente disporre i simboli degli alleli presenti nei gameti paterni e materni all'esterno dei due diversi lati di un quadrato, rispettivamente, e scrivere i genotipi degli individui risultanti in quattro quadrati più piccoli in cui si è diviso il primo quadrato).
Il fenotipo sarà normale se è presente almeno un gene normale A, e quindi i malati saranno il 25%, ossia in media un figlio su quattro. I soggetti omozigoti per l'allele A (genotipo AA) o eterozigoti (Aa) non manifesteranno il carattere, che in questo caso è l'albinismo, e saranno quindi normali. Però gli eterozigoti potranno trasmettere la malattia se si uniranno ad un altro dterozigote, o a maggior ragione a un malato.
Se la coppia avesse molti figli, questa percentuale potrebbe osservarsi con maggiore facilità. Tuttavia, poichè la combinazione mantiene un carattere di casualità, perché non vi è un meccanismo di "memoria" ma si tratta ogni volta di una estrazione a sorte, è anche possibile che i figli siano tutti sani, se non avviene mai l'incontro di un ovulo con genotipo a con uno spermio con genotipo a, o anche tutti malati, se sfortunatamente questo avviene per più volte di seguito. E' chiaro che più sono grandi i numeri osservati, più le percentuali si avvicineranno a quelle previste sulla base delle leggi di Mendel. Allo stesso modo, su un numero ristretto di lanci di una monetina, una certa faccia (ad es., "testa") può casualmente uscire più volte di seguito se eseguiamo pochi lanci, ma su un numero grande di lanci molto probabilmente uscirà con frequenza identica o molto simile a quella con cui si presenterà la faccia "croce". E' importante osservare allora che rimane impossibile fare previsioni affidabili sul caso singolo; si può solo quindi indicare una percentuale, una probabilità di avere testa o croce, che è identica ad ogni lancio ed è il 50%. Nel caso delle malattie recessive, analogamente, il rischio è di trasmettere la patologia è del 25% per ogni fecondazione avvenuta, qualunque sia stato l'esito delle fecondazioni precedenti. Solo se abbiamo la possibilità di osservare un numero molto grande di figli, o più generazioni della famiglia, o più famiglie con la stessa malattia, è probabile che globalmente il rapporto tra sani e malati si avvicini a quello atteso di 3:1.
La biologia
molecolare
ha permesso di chiarire l'esatto meccanismo che è alla base
dell'albinismo.
Nelle cellule esposte alla luce, la melanina è prodotta a
partire
dall'amminoacido tirosina. La conversione della tirosina in melanina
avviene
ad opera di un enzima, detto TIROSINASI. Si ricordi che gli enzimi sono
di solito catene polipeptidiche, che svolgono una certa funzione in
quanto
sono costitutite una determinata sequenza di amminoacidi; questa
sequenza
è specificata, attraverso le regole note come "Codice genetico",
dalla sequenza di basi del gene responsabile della produzione di quella
catena polipeptidica. Il gene per la Tirosinasi è localizzato
sul
cromosoma 11 (11q14, ossia quarta banda del primo gruppo di bande sul
braccio
lungo del cromosoma 11), e ciascuno ne possiede due copie: quella che
si
trova sul cromosoma 11 paterno e quella che si trova sul cromosoma 11
materno.
L'allele che causa l'albinismo è una versione mutata del gene
per
la tirosinasi, che porta alla sintesi di una Tirosinasi di sequenza
amminoacidica
sbagliata e incapace di convertire la tirosina in melanina. Se
l'individuo
ha almeno una copia corretta del gene per la tirosinasi, può
comunque
produrre melanina sufficiente; ma se riceve entrambe le copie errate
(l'allele
a), può produrre solo enzima di sequenza sbagliata e non
funzionante,
con conseguente assenza di conversione di tirosina in melanina. Il
risultato
finale è la mancanza di melanina solo nei soggetti omozigoti per
l'allele mutato a, i soli che presenteranno l'aspetto albino descritto.
Si osservi anche che il fenotipo è spiegato a sufficienza dalla
mutazione del gene della tirosinasi, indipendentemente dall'ambiente in
cui vive il soggetto, ad es. dalla sua esposizione alla luce, e
indipendentemente
dalla mutazione di altri geni. Si tratta quindi di una tipica malattia
monofattoriale.
2. EREDITA' AUTOSOMICA DOMINANTE
Consideriamo un albero genealoghco esemplificativo di una famiglia in cui vi è un soggetto affetto da nanismo, nella tipica forma di nanismo acondroplasico (acondroplasia, da "a", assenza, "plasia", sviluppo, "condro", cartilagine). Questa malattia è nota fin dall'antichità, e consiste in una alterazione dello sviluppo scheletrico per cui i soggetti sono di bassa statura, in media 131 cm gli uomini e 124 cm le donne, e presentano un aspetto caratteristico del volto, con prominenza frontale accentuata. La patologia va distinta da altre forme di nanismo, come ad es. quella causata da difetti della tiroide, che presentano caratteristiche cliniche e cause differenti.
Osserviamo che un genitore risulta affetto, e dei quattro figli sono due quelli colpiti dalla malattia. Sono ugualmente colpiti maschi e femmine.
Le
caratteristiche
principali e le relative deduzioni sono allora: l'eredità non
appare
legata al sesso, quindi è autosomica; vi è un malato in
tutte
le generazioni e appare colpito un numero elevato di soggetti, in
accordo
con il comportamento che ci si attende per un carattere dominante.
Si tratta in effetti
di una malattia con eredità autosomica dominante, e in questo
caso
l'allele A (indicato con la maiuscola perché dominante) è
quello mutato e che causa la malattia, mentre "a" sarà l'allele
normale. Scriveremo allora in questo modo i genotipi degli individui
che
si incrociano:
Aa x aa
Secondo quanto sopra osserwato per la meiosi, i gameti possibili saranno:
A, a a, a
Considerando le possibili combinazioni alla fecondazione, i genotipi dei figli potranno quindi essere, ciascuno con probabilità del 25% (1/4):
Aa Aa aa aa
Il fenotipo
sarà
patologico se è presente almeno un allele A mutato,
poiché
A presenta un effetto di dominanza sull'allele normale a. Pertanto i
malati
saranno il 50%, ossia (in media) la metà dei figli. Solo
soggetti
omozigoti per l'allele a (genotipo aa) non manifesteranno il carattere,
che in questo caso è il nanismo, e saranno quindi sani.
Nella realtà,
vale quanto esposto a proposito della eredità autosomica
dominante:
il rischio di avere un figlio malato è per questa coppia del
50%,
ma questo non esclude a priori che per caso i figli non possano essere,
ad es., tutti sani o tutti malati. Si osservi anche che nel caso della
acondroplasia l'allele mutato è letale in omozigosi, ossia il
genotipo AA determina gravi malformazioni scheletriche che portano a
morte l'individuo nel periodo neonatale per deficit respiratorio dovuto
alla piccolezza della gabbia toracica.
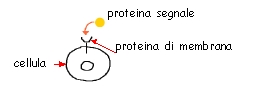
Solo nel 1994 la biologia molecolare ha permesso di chiarire l'esatto meccanismo che è alla base del nanismo. Il gene mutato nel nanismo si trova sul cromosoma 4 umano (4p16.3), e codifica per una proteina di membrana. Questa proteina di trova immersa nella membrana cellulare e funziona come un recettore, cioè un dispositivo in grado di captare un segnale.
In questo caso il
segnale
è una proteina che stimola le cellule connettivali a dividersi,
detta FGF ("Fibroblast growth factor",
letteralmente "fattore di
crescita
dei fibroblasti", anche se in realtà questo fattore
induce
la divisione cellulare anche nei condrociti, negli osteociti ed in
altre
cellule di tipo connettivale e non connettivale, e anche se in alcuni casi inibisce la proliferazione
cellulare anziché stimolarla). Vi sono diversi tipi
di FGF. Il gene del nanismo permette di produrre una delle varie
proteine
che funzionano come recettori per l'FGF, dette pertanto FGFR (ossia
"Recettore
per l'FGF"; nella acondroplasia è implicato il terzo tipo dei
diversi
recettori per gli FGF presenti nell'organismo, detto FGFR3
- fibroblast growth factor receptor 3).
Le cellule
che espongono questo recettore sono sensibili alla azione degli FGF,
perché
sono in grado di legarlo. Il legame del fattore di crescita al
recettore
avvia una serie di reazioni chimiche nel citoplasma, in cui sporge la
"coda"
del recettore.
L'allele
responsabile
del nanismo è nel 99% dei casi un allele con una sola base
cambiata
(sostituzione di un singolo nucleotide) per cui a livello del codone n.
380 (a partire dal codone d'inizio) la tripletta GGG assume la sequenza
AGG. Poiché secondo il codice genetico GGG codifica per
l'amminoacido
Glicina (G) e AGG per l'amminoacido Arginina (R), la proteina
recettoriale
mutata presenterà un amminoacido sbagliato nella posizione 380
della
catena polipeptidica, a livello della regione della proteina infilata
nella
membrana della cellula. Riferendosi alla proteina, la mutaziooe si
può
indicare anche con la sigla G380R,
che si legge "R al posto di G in
posizione
380". Questo unico cambiamento amminoacidico è però
sufficiente
ad alterare il Recettore: il recettore mutato acquisisce la
proprietà di attivarsi anche in assenza del fattore che lo lega
(ligando), e diventa più resistente alla degradazione nei
lisosomi. L'effetto complessivo è quindi quello di una amplificazione dei segnali mediati
da FGFR3.
Il gene per FGFR3 è espresso ad alti livelli negli abbozzi
cartilaginei preossei di tutte le ossa. E' stato dimostrato che FGFR3
è in realtà un regolatore
negativo della
proliferazione e del differenziamento dei condrociti ddlla
cartilagine di accrescimento, e che è attivato dalle mutazioni
presenti nei soggetti con acondroplasia. Queste mutazioni sono
da considerare del tipo con "guadagno di funzione" (gain of function),
perché il recettore diventa
più efficace nello
svolgimento del suo ruolo inibitorio. La funzione eccessiva del
recettore mutato impedisce pertanto un normale accrescimento lineare
dell'osso. Le cartilagini di accrescimento degli individui che portano
la mutazione G380R nel gene FGFR3 sono in effetti disorganizzate,
presentano un numero ridotto di cellule e mostrano una maturazione
aberrante dei condrociti. Basta una sola copia
del gene mutato, come negli eterozigoti Aa, per
causare
la malattia, perché in questi casi metà dei rebettori che
ogni cellula esporrà funzioneranno normalmente, ma
la restante
metà
funzionerà eccessivamente, per cui il risultato finale
sarà
comunque un eccesso di funzione.
Si osservi che
anche
in questo caso parliamo di una tipica malattia monofattoriale, il cui
il
fenotipo è spiegato a sufficienza dalla mutazione del gene
FGFR3,
indipendentemente dall'ambiente in cui vive il soggetto, ad es. dalla
sua
dieta, e indipendentemente dalla mutazione di altri geni.
3. EREDITA' LEGATA AL SESSO
Poiché il caso più frequente è quello di geni presenti sul cromosoma X, si parla anche di "eredità legata all'X" o "eredità diaginica", in quanto si trasmette attraverso (in greco "dià") la donna (in greco "ginaikòs"). Si trasmettono con questa modalità alcune importanti malattie genetiche umane, come la emofilia A e la distrofia muscolare.
Consideriamo come esempio classico l'albero genealogico della famiglia reale inglese, nella quale alcuni membri furono affetti a un certo punto dalla EMOFILIA (dal greco "philìa", amicizia, affinità e "haima", sangue, ossia: "tendenza a sanguinare, ad avere emorragie").
Questa malattia consiste in un grave difetto della coagulazione del sangue. Si nota, osservando un tipico albero genealogico, che sono colpiti solamente i maschi, mentre per il resto le caratteristiche del tipo di trasmissione appaiono simili a quelle descritte per le malattie autosomiche recessive.
Le basi
molecolari della malattia consistono nella mutazione del gene per il
FATTORE VIII
(fattore ottavo), una proteina importante per il processo di
coagulazione
normale. Il gene è localizzato sul cromosoma X, per cui,
indicando
con A l'allele normale del gene per il fattore VIII e con a l'allele
mutato,
che produce una proteina non funzionante, si deduce che la donna
può
essere, se consideriamo l'allele mutato, o eterozigote (Aa) o omozigote
(aa). Viceversa, l'uomo possiede una sola copia del cromosoma X, per
cui
avrà una sola copia del gene per il fattore VIII, o normale (A)
o mutata (a). Questa particolare situazione si verifica per tutti i
geni
del cromosoma X nel maschio, che viene detto
EMIZIGOTE per questi
geni; in effetti non si può parlare di omozigosi o eterozigosi,
termini riferiti alla identità o diversità delle due
copie
alleliche del gene, se normalmente in questo caso deve essere presente
una sola copia.
La situazione più frequente in assoluto è quella di una donna portatrice che sposa un uomo non emofilico, per cui secondo lo schema mendeliano avremo questi genotipi nei figli: (indichiamo con Xm il cromosoma X che porta il gene mutato): 25% XX (femmine normali), 25% XXm (femmine sane ma portatrici), 25% Xy (maschi sani), 25% Xmy (maschi malati di emofilia).
La DISTROFIA
MUSCOLARE
segue un analogo schema di trasmissione. Il gene mutato si trova sul
cromosoma
X e codifica per la distrofina, una proteina coinvolta nella
contrazione
muscolare. I bambini affetti presentano gravi disturbi del movimento e
non raggiungono l'età adulta per insufficienza dei muscoli
respiratori.
Attualmente, una
diagnosi
precoce entro il secondo-terzo anno di vita, un sollecito e corretto
trattamento
riabilitativo e un adeguato supporto farmacologico possono aumentare la
aspettativa di vita media dei pazienti dai 20 anni del passato anche
fino
a 30 anni.
4. NOTE CONCLUSIVE: PENETRANZA E CONSANGUINEITA'
La trasmissione
della
malattia si è fatta equivalere a quella dell'allele mutato, come
in effetti spesso avviene per le malattie monofattoriali in cui la
componente
genetica spiega praticamente tutta la malattia. Tuttavia può
anche
accadere che una certa percentuale di soggetti possieda un genotipo che
dovrebbe causare la malattia, ma risulta sano.
La PENETRANZA del
carattere è la percentuale di soggetti in cui il genotipo mutato
si manifesta.
Ad esempio, la
acondroplasia
è una malattia con penetranza del 100% (penetranza completa),
perché
tutti i soggetti con la mutazione del gene FGFR3 di fatto manifestano
la
malattia. Se ad esempio su 100 soggetti con la mutazione fossero malati
solo 70, diremmo che la penetranza è del 70%.
Il fenomeno della
penetranza variabile indica l'esistenza di altri fattori (altri geni o
fattori ambientali) che collaborano, seppure in minor parte, alla
manifestazione
della malattia.
Una osservazione finale va fatta considerando che le malattie recessive (autosomiche o legate all'X) sono rare nella popolazione, perché è improbabile che un portatore si unisca ad un altro portatore. Tuttavia questa probabilità aumenta se due soggetti di una famiglia in cui è presente l'allele mutato si uniscono. Da tempo si è osservato che il rischio di malattie genetiche è più alto nei matrimoni tra consanguinei ("che hanno lo stesso sangue", ossia parenti), ed è tanto più alto quanto maggiore è il grado di CONSANGUINEITA'. Un rischio piuttosto elevato è presente nei matrimoni tra primi cugini, e nelle popolazioni con maggiore frequenza di matrimoni tra consanguinei, come quelle insulari, per motivi geografici, o quelle che valorizzano l'unione tra parenti per motivi storici e culturali (famiglie regnanti, alcune tribù, etc).
GLOSSARIO DI
RIEPILOGO
GENE = porzione
di
DNA che codifica per un prodotto (una catena polipeptidica).
LOCUS = posizione
del gene sul cromosoma.
ALLELE = una forma
alternativa di un gene.
OMOZIGOTE = un
individuo
che presenta, in un dato locus, due alleli uguali.
ETEROZIGOTE = un
individuo
che presenta, in un dato locus, due alleli diversi.
CARATTERE
MONOFATTORIALE
= il carattere è controllato da un solo fattore, ossia da un
singolo
gene.