Un cromosoma
è una molecola di DNA che contiene l'informazione genetica di un
organismo.
Nell'uomo i cromosomi sono 46 molecole lineari di DNA a doppia elica.
23
molecole derivano dalla madre, 23 dal padre; per questo si dice che il
corredo genetico umano è composto da 23 coppie di cromosomi.
Le molecole di
DNA
sono molto lunghe: se si considera che le coppie di basi nel genoma
umano
sono 6,4 miliardi, in media ogni cromosoma è costitutito da 139
milioni
di coppie di basi, che occupano circa 4,7 cm in lunghezza, quindi la
lunghezza
totale dei cromosomi umani di è approssimativamente di due metri
in ogni singola cellula. Per poter essere contenute in uno spazio
ridotto
come il nucleo cellulare, le molecole di DNA devono allora assumere una
struttura compatta. A tale scopo la molecola di DNA a doppia elica si
avvolge
attorno a delle proteine basiche chiamate ISTONI. Esistono quattro tipi
di istoni:
H1 (ricchi in lisina)
H2 (istoni intermedi)
H2b
H3 (ricchi in
arginina)
H4
Gli istoni H2, H2b,
H3 e H4 si dispongono a formare quattro spicchi di una torta, impilata
su un'altra uguale in una struttura detta ottamero ("formato da otto
parti").
Un tratto di DNA di circa 146 basi compie un doppio giro attorno
all'ottamero.
La molecola si avvolge ulteriormente su se stessa per diverse volte, in
un processo detto spiralizzazione, aumentando cosi' di spessore mentre
si riduce di lunghezza.
La massima spiralizzazione si ha durante la divisione cellulare (mitosi), mentre quando la cellula non è in mitosi il DNA viene svolto, in particolare nelle regioni che contengono geni che vengono trascritti.
Durante la divisione cellulare ogni molecola di DNA risulta quindi accuratamente ripiegata attorno ad un'impalcatura composta di proteine, formando un bastoncello visibile al microscopio ottico e intensamente colorabile: per questo si dette il nome a ciascun bastoncello di cromosoma, ossia "corpuscolo colorato", dal greco "croma" (colore) e "soma" (corpo). Poichè la cellula in mitosi ha replicato tutto il suo DNA, ciascun bastoncino si trova associato a un bastoncino che ne è la copia, avendo la stessa sequenza di basi. Per questo, quando possiamo concretamente osservare nel modo migliore i cromosomi, e cioè alla metafase, i cromosomi appaiono a forma di X: due bastoncini attaccati tra loro a livello della zona centrale. In questo stadio ogni cromosoma è quindi composto da due parti dette CROMATIDI. Poiché la molecola di DNA che compone un cromatide di un cromosoma ha la stessa sequenza della molecola di DNA che costituisce l'altro, i due cromatidi di uno stesso cromosoma sono anche detti "cromatidi fratelli".
La condensazione dei cromosomi ne facilita una ripartizione ordinata, senza pericolo di aggrovigliamento tra molecole di DNA diverse. La cellula quindi prima duplica tutto il DNA nucleare, poi lo divide tra le due cellule figlie. Pertanto solo durante le prime fasi della divisione cellulare il corredo dei cromosomi apparira' come un insieme di 46 elementi a forma di "X" (due cromatidi uniti a livello del centromero, per un totale di 92 cromatidi), mentre alla anafase i due cromatidi si separeranno e appariranno come due bastoncini singoli a forma di "I". Nel periodo che intercorre tra una divisione e l'altra il materiale genetico normale è in effetti composto, per ogni cellula somatica, da 46 cromatidi singoli, che tuttavia noi non visualizziamo perché queste 46 molecole di DNA sono despiralizzate in filamenti sottilissimi a formare la cromatina.
Si noti che la cellula, dividendosi, dimezza la quantita' del citoplasma, che viene diviso fra le due cellule figlie, ma l'informazione genetica, essendo stata prima raddoppiata e poi divisa a metà tra le due cellule figlie, rimane identica. Solo nel caso delle cellule germinali mature vi è un effettivo dimezzamento del materiale genetico, in seguito alla meiosi, perché ciascun gamete possiede 23 cromosomi in totale.
Analisi del cariotipo umano
Il cariotipo
di una cellula è il suo corredo, o assetto, cromosomico;
la parola indica il "tipo" di assetto che si osserva nel nucleo, in
greco
"carion". Il cariotipo è caratteristico di ciascuna specie.
La citogenetica
è una branca della genetica che ha per oggetto lo studio
morfologico
della struttura, della topologia e della funzione dei cromosomi.
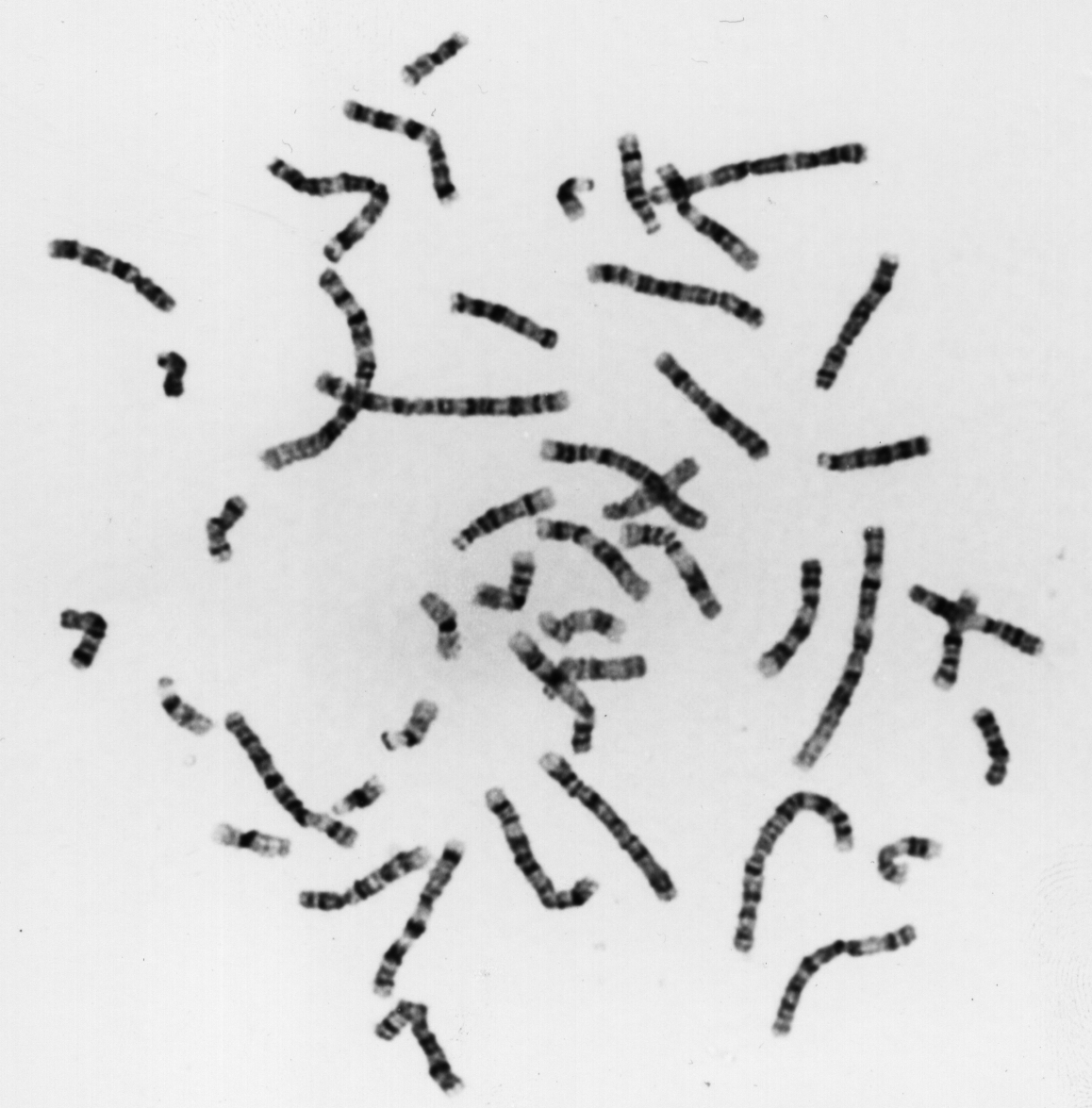
Il momento più adatto per osservare e contare i cromosomi si presenta durante la mitosi.
SCHEMA
http://www.accessexcellence.org/AB/GG/human.html
Negli organismi
adulti
la divisione cellulare è un fenomeno costante solo in alcuni
tessuti,
le cui cellule a vita breve devono essere continuamente sostituite. Le
cellule soggette a rinnovamento continuo sono, tra quelle somatiche, le
cellule del sangue, dell'epidermide e della mucosa intestinale, e, tra
quelle germinali, gli spermatozoi.
La modalità
più semplice dal punto di vista clinico è quella di
studiare
il cariotipo a partire dalle cellule ottenute mediante un normale
prelievo
di sangue. Le cellule del sangue adatte all'osservazione del cariotipo
dovranno avere necessariamente il nucleo (e quindi non potranno essere
i globuli rossi, o gli elementi piastrinici) e dovranno poter essere
ancora
capaci di entrare in mitosi, come lo sono i linfociti, uno dei tipi di
globuli bianchi.
La procedura
pertanto
può essere così sinteticamente descritta: si separano dal
sangue in provetta le cellule linfocitarie e si pongono in coltura con
un terreno nutritivo in un adatto incubatore, alla temperatura di
37°C.
In queste condizioni le cellule possono continuare a vivere all'esterno
dell'organismo. Si induce quindi la divisione cellulare aggiungendo una
sostanza detta "fitoemoagglutinina", in inglese
"PhitoHemoAgglutinin"
(sigla:
PHA). La PHA è una lectina estratta dalle piante
(fito) che è in grado di provocare la agglutinazione dei globuli
rossi. Inoltre, i linfociti stimolati dalla PHA entrano in mitosi e
iniziano
a dividersi.
A questo punto viene
aggiunta la colchicina, un composto tossico vegetale che si
comporta
come un veleno per i microtubuli, bloccandone l'azione. La mitosi
quindi
viene bloccata in tutte le cellule allo stadio di metafase, con 46
cromosomi
ben spiralizzati, tutti composti da due cromatidi e allineati sul piano
centrale della cellula ("equatore"), perché i microtubuli non
riescono
più a separare e a trascinare i cromatidi, con la conseguente
impossibilità
di iniziare l'anafase.
Rimane ora il
problema
di allontanare tra loro i cromosomi, in quanto seppure ben visibili al
microscopio risultano affollati nella stessa regione e sono quindi
difficili
da osservare e contare. Per questo le cellule vengono immerse in una soluzione
ipotonica, cioè a minor concentrazione salina di quella
presente
all'interno del citoplasma. Poiché i sali non hanno passaggio
libero
attraverso le complesse e selettive membrane cellulari, sarà
l'acqua
a spostarsi verso l'interno delle cellule, per permettere il
raggiungimento
di un equilibrio tra la concentrazione salina all'esterno della cellula
e la concentrazione salina, più elevata, presente all'interno
delle
cellule; questo fenomeno è detto "osmosi". In definitiva, le
cellule
assorbono acqua, sino a gonfiarsi e poi scoppiare. Si noti che se al
contrario
la soluzione fosse stata ipertonica, la cellula avrebbe perso acqua,
raggrinzendosi.
La rottura della cellula causa lo spargimento nei dintorni dei cromosomi. Ora è possibile osservarli su un vetrino al microscopio, e fotografarli. Dalla foto di un gruppo di cromosomi vengono ritagliate le sagome di ciascun cromosoma, e in questo modo è possibile disporre l'immagine di ogni cromosoma ordinatamente su un foglio di carta, eseguendo una sorta di collage. I cromosomi vengono disposti in ordine decrescente rispetto alle dimensioni, dal più grande al più piccolo, e si ha cura di mettere vicini, a coppie, i due cromosomi omologhi. Al termine, potremo osservare che sono presenti 22 coppie di cromosomi omologhi, numerate da 1 a 22, per un totale di 44 cromosomi (ad es.: due cromosomi 1, due cromosomi 2, due cromosomi 3, e così via fino a 22; ciascun cromosoma è composto di due cromatidi perché non abbiamo permesso l'ultimazione della mitosi). Si noti che in effetti il cromosoma più piccolo è il 21, non il 22, per un errore iniziale nella valutazione delle dimensioni degli autosomi.
I due rimanenti
cromosomi
formano una coppia solo nella donna, mentre nell'uomo sono molto
diversi
tra loro, essendo uno molto più piccolo dell'altro. Per questa
ragione
sono detti "cromosomi del sesso" o "eterocromosomi" (da
"etero",
diverso), mentre i cromosomi non sessuali sono detti "autosomi"
(da "autos", uguale, perché sono tutti presenti in coppie
composte
da due elementi simili tra loro). I cromosomi del sesso sono di due
tipi,
detti "X" e "Y" per la somiglianza della loro forma a queste due
lettere
dell'alfabeto. Il cromosoma X è piuttosto grande, di dimensioni
simili a quelle del cromosoma 6, mentre il cromosoma Y è molto
piccolo
e contiene pochi geni. Il corredo cromosomico femminile comprende due
cromosomi
X, mentre nel cariotipo maschile si osserva un cromosoma X e un
cromosoma
Y.
In sintesi, il
cariotipo
degli individui della specie Homo sapiens si indica dichiarando il
numero
totale dei cromosomi, e quindi indicando, dopo una virgola, il tipo di
cromosomi del sesso.
Per una donna avremo
quindi:
46,XX
Per un uomo:
46,XY
La differenza genetica tra uomo e donna solo a livello dei cromosomi del sesso fa supporre, come è vero, che questi cromosomi contengano informazioni necessarie al differenziamento del sesso.
Forma dei cromosomi
Una osservazione
più
attenta dei cromosomi disposti ordinatamente per dimensione, come sopra
indicato, permette di rilevare anche differenze di forma, dovute alla
diversa
posizione del centromero.
Ogni cromatide
è
idealmente diviso in due parti dal centromero. La regione più
corta è detta "braccio corto", o "p" (dal francese "petit",
piccolo),
e la regione più lunga è detta "braccio lungo".
Ciascun
cromosoma, essendo composto di due cromatidi, presenta quattro braccia,
due corte uguali tra loro e due lunghe uguali tra loro.
Nei cromosomi
"metacentrici"
il centromero si trova all'incirca a metà della lunghezza dei
cromatidi;
mentre in quelli
"submetacentrici"
si trova leggermente spostato verso una estremità.
In alcuni cromosomi,
infine, il centromero è posto vicino ad una delle due
estremità
del cromosoma stesso; questi cromosomi, cioè le coppie 13-14-15
(gruppo D), 21-22 (gruppo G), e il cromosoma Y, sono detti
"acrocentrici",
dal greco "acros" (sommità), e assomigliano più a una "Y"
che a una "X". I cromosomi telocentrici (con centromero in posizione
terminale)
non si osservano nell'uomo.
La differenza di
lunghezza
delle due braccia del cromatide è ovviamente particolarmente
evidente
nei cromosomi acrocentrici, in cui il braccio p si estende per molto
poco
al di là del centromero.
GRUPPI DI
CROMOSOMI
UMANI
http://homepages.uel.ac.uk/V.K.Sieber/human.htm
Bandeggio dei cromosomi umani
Sebbene
l'osservazione
delle differenze di dimensioni e di quelle di forma potrebbe essere
sufficiente
a stabilire l'identità di ciascun cromosoma sul cariotipo,
esistono
anche cromosomi diversi non facilmente distinguibili tra loro solo
sulla
base di questi due criteri.
Vengono quindi
applicate
delle colorazioni artificiali che permettono di evidenziare differenze
più fini tra i cromosomi e di stabilire inequivocabilmente la
coppia
di appartenenza per ciascun cromosoma.
Questi coloranti
tipicamente
tingono solo alcune regioni del cromosoma, lasciando le altre chiare,
generando
così un assetto "a striscie" caratteristico di ciascuna coppia
di
omologhi. Queste "fasce" vengono chiamate BANDE e l'operazione di
colorazione
che ha il fine di ottenerle si chiama
bandeggio.
La chinacrina genera
bande scure dette Q (dall'inglese "Quinacrine"), mentre il colorante
Giemsa
evidenzia bande dette "G". Si è scoperto che le regioni che
appaiono
scure con questi coloranti sono ricche di basi azotate A e T, mentre le
zone cromosomiche la cui sequenza presenta maggiore abbondanza di G e C
rimangono non colorate.
BANDE
http://www.pathology.washington.edu/research/cytopages/spreads/human_g.gif
http://www.emc.maricopa.edu/faculty/farabee/BIOBK/Human_46,XY.gif
http://www.tokyo-med.ac.jp/genet/cki-e.htm
Ogni banda ha una
sua
nomenclatura, che permette di ricavarne la posizione, come su una
mappa.
Se diremo che un gene, ad es., è localizzato in 6p21.3,
interpreteremo
ogni dato da sinistra a destra in questo modo: cromosoma, braccio,
regione,
banda sottobanda. Quindi:
6
- è il numero del cromosoma (cromosoma 6)
p
-
ci riferiamo al braccio più corto del cromosoma
2
-
il secondo gruppo di bande (regione) visibile sul braccio p, a partire
dal centromero
1
-
la prima banda all'interno di questo gruppo, contando dal centromero
verso
il telomero
3
-
il numero preceduto da un punto indica una "sottobanda", una banda
sottile
visibile all'interno di una banda più spessa; in questo caso la
terza sottobanda 1, sempre contata a partire dal centromero.
In altre parole, la posizione indicata è la terza sottobanda della prima banda osservabile sul secondo gruppo di bande localizzato sul braccio corto del cromosoma 6.
Per avere un
punto
di riferimento, si possono consultare schemi delle bande presenti in
ciascun
cromosoma umano, approvati a livello internazionale. Con tre tipi di
tecniche
diverse, si possono ottenere nell'intero cariotipo umano 400, 550
oppure
850 bande, rispettivamente. Gli "ideogrammi" sono rappresentazioni
schematiche
del bandeggio di ciascun cromosoma ottenibile con ciascun tipo di
metodica.
La rappresentazione
del corredo cromosomico come serie di cromosomi bandeggiati è
detta
cariogramma.
IDEOGRAMMI DEI
CROMOSOMI
UMANI
http://www.pathology.washington.edu/research/cytopages/idiograms/human/
Si noti che un difetto su un braccio di un cromatide di un cromosoma, è presente anche sul braccio corrispondente dell'altro cromatide, che ne è la copia. Una banda può contenere decine o centinaia di geni; la posizione del gene sul cromosoma si chiama locus (plurale: loci).
MAPPA FISICA DEI
CROMOSOMI
UMANI
http://www.ncbi.nlm.nih.gov/mapview/map_search.cgi?chr=hum_chr.inf&query
Poichè la più piccola alterazione di un cromosoma visibile mediante l'analisi del cariotipo standard è quella di una banda o sottobanda, che in media comprende un tratto di DNA della lunghezza di 4 milioni di basi, ne consegue che le alterazioni visibili osservando il cariotipo sono quelle di grande estensione, come le mutazioni genomiche o cromosomiche, mentre mutazioni che riguardano poche basi non si potranno evidenziare con questa metodica. Metodiche più recenti permettono di ottenere risoluzioni molto superiori.
In conclusione,
l'analisi
del cariotipo permette di stabilire:
- la specie
dell'individuo
- il sesso
dell'individuo
- la presenza di
eventuali
alterazioni
del numero e del tipo di cromosomi
- la presenza di
eventuali
alterazioni
della struttura dei cromosomi
Mitosi, meiosi
e
fecondazione:
significato
genetico
La divisione cellulare comporta la replicazione delle informazioni genetiche della cellula madre e la sua ridistribuzione nelle cellule figlie.
La mitosi
è
la divisione delle cellule somatiche, e il trattamento
dell'informazione
genetica è mirato semplicemente alla conservazione
dell'informazione
genetica nelle due cellule figlie.
Dal punto di vista
genetico, riassumiamo gli aspetti informazionali di una cellula umana
("Cellula
madre") che si divide generando due cellule figlie ("Cellula figlia 1"
e "Cellula figlia 2"). I parametri che seguiremo sono: c ("DNA
content",
il contenuto in DNA, relativo a quello di una cellula aploide); il
numero
di cromosomi, dove n=23, numero aploide, nella specie umana; il tipo di
cromosomi (composto di una sola molecola a doppia elica, ossia
"monocromatidico",
oppure di due doppie eliche identiche, ossia "dicromatidico", con due
cromatidi
fratelli); la dimensione in Gbp (miliardi di coppie di basi);
| Cellula | Contenuto in DNA | Numero di cromosomi | Tipo di cromosomi | Dimensione del genoma | |
| Cellula madre | 2c | 2n = 46 | Monocromatidico | 6,4 Gbp | |
| Cellula madre dopo la fase S (replica del DNA) | 4c | 2n = 46 | Dicromatidico | 12,8 Gbp | |
| Cellula figlia 1 | 2c | 2n = 46 | Monocromatidico | 6,4 Gbp | |
| Cellula figlia 2 | 2c | 2n = 46 | Monocromatidico | 6,4 Gbp | |
Il significato genetico è quindi quello di ricostituire nelle cellule figlie la stessa informazione, qualitativa e quantitativa, della cellula madre. Se ciò fosse sempre vero, un individuo multicellulare sarebbe composto di cellule tutte geneticamente identiche al 100% tra loro. In realtà, per effetto di mutazioni puntiformi originatesi durante la replicazione del DNA e sfuggite ai sistemi di riparazione, si stima che circa 10-9 basi per genoma siano diverse (6 basi in ogni cellula), con un accumulo progressivo ad ogni mitosi, per cui le cellule somatiche dell'adulto possono avere alcune decine di geni mutati rispetto a quelli presenti nello zigote, tollerati perché non influenti sulle funzioni dello specifico tipo cellulare differenziato.
La meiosi
è
la divisione cellulare tipica delle cellule germinali negli individui a
riproduzione sessuata, e presenta due particolarissimi fenomeni nel
trattamento
delle informazioni: il dimezzamento del genoma, e la
generazione
di nuove sequenze cromosomiche rispetto a quelle originarie.
Il dimezzamento del
numero dei cromosomi nella cellula sessuale matura (gamete)
permette
di mantenere costante tale numero nella specie, in quanto in seguito a
unione dei gameti dei due sessi, entrambi con numero aploide di
cromosomi,
si ricostituisce il doppio corredo cromosomico (diploide) tipico delle
cellule somatiche, che possiedono una versione paterna e una materna di
ciascun cromosoma. La riduzione del materiale genetico avviene in due
fasi:
nella prima divisione cellulare (meiosi I) il precursore dei
gameti
replica il DNA, ma durante la divisione cellulare i cromatidi non si
separano,
per cui ciascuna cellula riceve 23 cromosomi dicromatidici, ossia un
solo
cromosoma per ciascuna coppia di omologhi. Segue una seconda divisione
cellulare (meiosi II) non preceduta da replicazione del DNA,
durante
la quale i due cromatidi dei 23 cromosomi di ciascuna cellula si
separano,
ottenendosi alla fine due cellule figlie (da ciascuna delle cellule
prodotte
dalla meiosi I) con 23 cromosomi monocromatidici, patrimonio che
corrisponde
al 50% di quello delle cellule somatiche.
| Cellula | Contenuto in DNA | Numero di cromosomi | Tipo di cromosomi | Dimensione del genoma |
| Spermatogonio | 2c | 2n = 46 | Monocromatidico | 6,4 Gbp |
| Spermatocita di I ordine
dopo la fase S (replicazione del DNA) |
4c | 2n = 46 | Dicromatidico | 12,8 Gbp |
| Meiosi I | ||||
| Spermatocita II ordine (prima cellula figlia) |
2c | n = 23 | Dicromatidico | 6,4 Gbp |
| Spermatocita II ordine (seconda cellula figlia) |
2c | n = 23 | Dicromatidico | 6,4 Gbp |
| Meiosi II | ||||
| Spermatide (prima cellula figlia) |
c | n = 23 | Monocromatidico | 3,2 Gbp |
| Spermatide (seconda cellula figlia) |
c | n = 23 | Monocromatidico | 3,2 Gbp |
Inoltre, il dimezzamento del materiale genetico è associato anche alla prima fonte di variabilità della informazione genetica nei gameti: l'assortimento indipendente, ossia la distribuzione casuale di ciascun membro di una coppia di cromosomi omologhi in una delle due cellule figlie all'atto della anafase I. Ad esempio, prendendo in esame la coppia di cromosomi 1, il cromosoma 1 paterno potrebbe segregare nella cellula figlia 1, e il corrispettivo materno nella cellula figlia 2; il contrario potrebbe avvenire casualmente per la coppia dei cromosomi 2, e così via. Se si considerano in H. sapiens le 23 coppie dei cromosomi, i membri di ciascuna delle quali vengono distribuiti a caso tra le due cellule figlie, si hanno 223 combinazioni possibili, ossia più di 8 milioni di gameti diversi solo per effetto dell'assortimento indipendente durante il dimezzamento del genoma.
Sebbene tale variabilità sia di per sé enorme, va anche sottolineato che durante la meiosi I, durante la profase, i rispettivi cromosomi dicromatidici omologhi si sovrappongono (appaiamento degli omologhi), e vanno incontro al crossing-over, un meccanismo per cui due dei cromatidi sovrapposti vengono tagliati, e i segmenti generati dal taglio su ciascun cromatide si scambiano la posizione, finendo per trovarsi sull'altro membro della copia di cromosomi. In questo modo, si formano due nuove doppie eliche, ciascuna composta di una parte di origine paterna e una parte di origine materna. Il processo può essere immaginato come la creazione di nuovo mazzo di carte (i geni) a partire da due mazzi originariamente omogenei, o come l'assemblaggio di un nuovo programma mescolando le istruzioni di due programmi originari. Poiché il numero di punti di scambio e la loro posizione sono casuali, il numero possibile di nuove sequenze di DNA che ciascuna coppia di cromosomi appaiati può generare dopo il crossing-over è inconcepibilmente alto, ed è praticamente impossibile che una singola cellula sessuale di un individuo ripeta la stessa configurazione di crossing-over per tutte e 23 le coppie di cromosomi che si è verificata in un'altra cellula.
Nella specie umana, il numero di scambi per ogni cellula in cui avviene il crossing over è in media di 27 nel maschio (intervallo: 17-37, deviazione standard: ±7) e di 42 nella femmina (intervallo: 28-61, deviazione standard: ±4). Nella gametogenesi femminile si ha quindi costantemente un maggior tasso di crossing-over, per ragioni non chiare.
è importante notare come l'avvenimento di almeno uno scambio per ciascuna coppia di cromosomi omologhi sembri essere fondamentale per la realizzazione di una meiosi corretta: l'assenza di crossing over favorisce una segregazione anomala dei due cromosomi appaiati, che possono non separarsi dando origine a mutazioni cromosomiche di numero (una cellula riceve entrambi i cromosomi omologhi, e l'altra nessuno).
La fecondazione
segna l'inizio di un nuovo organismo, ed è il processo per cui due
cellule germinali si fondono insieme per creare un nuovo individuo con
un programma genetico derivato da entrambi i genitori. Alla
fecondazione,
una ulteriore (terza) fonte di variabilità genetica è
generata
dall'unione casuale di uno dei gameti paterni con uno dei gameti
materni.
In un episodio di fecondazione umana, sono in gioco circa 150 milioni
di
gameti maschili (spermatozoi) e uno tra i circa 400.000 gameti
femminili
(ovociti).
Se il significato
genetico della fecondazione è quello della ricostituzione di un
programma genetico completo, in grado di dirigere lo sviluppo del nuovo
individuo, si pone il problema della corretezza dei due singoli
programmi
genetici generati dai processi precedenti, e anche quello della
compatibilità
tra di due programmi stessi, perché specifiche combinazioni di
alleli
paterni e materni potrebbero non essere compatibili con uno sviluppo
normale.
Il controllo della qualità dei programmi genetici è
affidato
a meccanismi biologici di selezione naturale: gameti che portano gravi
mutazioni non sopravvivono, o sono svantaggiati durante la
competizione
per il processo di fecondazione. Inoltre, se lo squilibrio si forma
solo
nello zigote (o "one-cell embryo"), ed è grave, l'embrione
può
andare incontro ad aborto spontaneo.
In sintesi, i programmi genetici di ciascun genitore vengono dimezzati, rimescolati (mediante il crossing -over e l'assortimento casuale) e riuniti nella nuova cellula che si forma in seguito alla fecondazione. è quindi al momento della selezione del nuovo programma genetico, che avviene al momento dell'ingresso di quel determinato spermatozoo in un particolare ovocita, che risultano stabilite le caratteristiche ereditarie individuali.
Cromosomi,
Mitosi e Meiosi:
I video sono in formato .mov (Quick
Time e compatibili)
http://www.whfreeman.com/lodish4e/con_index.htm?09 (fare click su "Animations" e poi su "Three-Dimensional Packing of Nuclear Chromosomes")
http://academy.d20.co.edu/kadets/lundberg/DNA_animations/mitosis.mov
http://academy.d20.co.edu/kadets/lundberg/DNA_animations/mitosis1.mov
http://www.bio.davidson.edu/misc/movies/MEIOSIS.MOV
{kind=link}
{kind=link}